Пили — Википедия
Материал из Википедии — свободной энциклопедии
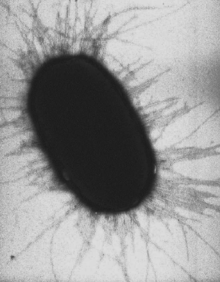 Клетки E. coli с многочисленными пилями
Клетки E. coli с многочисленными пилямиПили́, или фи́мбрии, или ворси́нки[1] — нитевидные белковые структуры, расположенные на поверхности клеток многих бактерий. Размер пилей варьирует от долей мкм до более чем 20 мкм в длину и 2—11 нм в диаметре. Пили участвуют в передаче генетического материала между бактериальными клетками (конъюгация), прикреплении бактерий к субстрату и другим клеткам, отвечают за адаптацию организмов, служат местами прикрепления многих бактериофагов.
Пили состоят из одного или нескольких типов спирально уложенных белковых молекул, которые называют пилинами[en] или фимбринами. Часто на концах пилей присутствуют особые прикрепительные субъединицы (адгезины), но иногда весь пиль обладает адгезивными свойствами. Структурно пили могут быть от тонких нитевидных образований до толстых палочкообразных структур с осевыми отверстиями. Очень тонкие пили (диаметром менее 2 нм) называют «кудряшками», они сливаются в пушистую липкую массу на поверхности клеток, которая обеспечивает их агрегацию. Пили-кудряшки являются амилоидными
Основная функция большинства пилей заключается в обеспечении правильного позиционирования особых молекул, обеспечивающих адгезию клеток. Взаимодействие бактерий с другими про- и эукариотическими клетками, обеспечиваемое пилями, является важным этапом заселения эпителия и проникновения патогенных бактерий в клетки организма хозяина, образования биоплёнок и переноса генетического материала при конъюгации. Некоторые пили задействованы в движении бактериальных клеток. Пили иногда служат рецепторами бактериофагов. Например, при инфекции мочевыводящих путей [en], вызванной патогенными штаммами Escherichia coli, клетки прикрепляются к эпителию мочевого пузыря с помощью пилей, на конце которых находятся молекулы белка-адгезина FimH, который взаимодействует с остатками маннозы на поверхности эпителиальных клеток. Благодаря прикреплению к эпителию клетки бактерии не выводятся из организма с мочой. Пили являются важными факторами вирулентности[en] многих кишечных бактерий, таких как Salmonella enterica, энтеропатогенные штаммы E. coli[en], Vibrio cholerae. Прикрепление пилей к эукариотическим клеткам может запускать в них определённые сигнальные каскады. Например, прикрепление пилей Neisseria к эпителиальным клеткам вызывает в последних выход ионов кальция в цитоплазму, при этом кальций является важным вторичным посредником в передаче сигнала в эукариотических клетках
В образовании пилей, помимо самих белков-пилинов, участвуют дополнительные белки, способствующие правильной сборке. У грамотрицательных бактерий они должны пройти через клеточную мембрану, периплазматическое пространство и наружную мембрану. Ключевую роль в сборке пилей играет периплазматический шаперон PapD и транспортный белок внешней мембраны, известный как Usher-белок. PapD связывается с пилинами, вышедшими в периплазму, и доставляет их к Usher-белку, после чего высвобождается назад в периплазму. Доставленные к Usher-белку пилины приобретают активированную конформацию и далее собираются в пили[6].
Пили-кудряшки[en] образуются по специфическому, тщательно регулируемому механизму. У E. coli в их сборке задействованы белковые продукты оперонов csgBA и csgDEFG. Фибрилла кудряшки образована белком CsgA, который, наряду с минорным белком CsgB, участвует в фазе нуклеации фибриллы. Когда нуклеация пройдена и начинается рост ворсинки, CsgB включается в неё. Фибриллы CsgA очень устойчивы и разрушаются лишь под действием 75 %-ной муравьиной кислоты. Оперон csgDEFG кодирует транскрипционный фактор CsgD и три предполагаемых фактора сборки пилей [2].
Традиционно пили подразделяют на 4 типа.
Некоторые учёные подразделяют пили на пили главного типа и альтернативного типа. Пили главного типа включают пили типов I—III по традиционной классификации. Они располагаются по всей поверхности клетки и прикрепляются непосредственно к её наружной поверхности. Пили главного типа выполняют только адгезионные функции. Пили альтернативного типа (тип IV по традиционной классификации) образуются в количестве 1—2 на клетку и располагаются на одном или двух её полюсах. Они, подобно жгутикам, имеют базальное тело и заякорены в клеточной мембране, а потому насквозь проходят периплазму и наружную мембрану. Пили альтернативного типа участвуют не только в адгезии. Они могут быстро разбираться от основания, при этом дистальные их концы остаются прикреплёнными к субстрату или другой клетке. За счёт этого одна клетка может подтягиваться ближе к другой, что особенно важно при конъюгации
- ↑ Нетрусов, Котова, 2012, с. 54.
- ↑ 1 2 Кассимерис, Лингаппа, Плоппер, 2016, с. 940.
- ↑ Нетрусов, Котова, 2012, с. 54—56.
- ↑ Нетрусов, Котова, 2012, с. 56—58.
- ↑ Кассимерис, Лингаппа, Плоппер, 2016, с. 939.
- ↑ Кассимерис, Лингаппа, Плоппер, 2016, с. 939—940.
- ↑ Пиневич, 2006, с. 338.
- ↑ Пиневич, 2006, с. 338—340.
- Пиневич А. В. Микробиология. Биология прокариотов: в 3 т. — СПб.: Издательство С.-Петербургского университета, 2006. — Т. I. — 352 с. — ISBN 5-288-04057-5.
- Нетрусов А. И., Котова И. Б. Микробиология. — 4-е изд., перераб. и доп. — М. : Издательский центр «Академия», 2012. — 384 с. — ISBN 978-5-7695-7979-0.
- Современная микробиология / Под ред. Й. Ленгелера, Г. Древса, Г. Шлегеля. — М.: Мир, 2005. — Т. 1. — 654 с.
Пили
Пили
Ворсинки или пили (фимбрии от англ. fimbria – бахрома), – тонкие полые нити белковой природы, более тонкие и короткие (3-10 нм х 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Они обладают антигенной активностью. По своему функциональному назначению пили подразделяются на несколько типов.
Пили 1-го типа, или общего типа – common pili – пили, ответственные за адгезию, т.е. за прикрепление бактерий к поражаемой клетке. Они начинаются на ЦПМ и пронизывают клеточную стенку. Их количество велико – от нескольких сотен до нескольких тысяч на одну бактериальную клетку. Бактериальные и эукариотические клетки заряжены отрицательно, но поверхностные микроворсинки снижают заряд бактерии и уменьшают электростатические силы отталкивания. Кроме того, увеличение площади поверхности бактериальной клетки дает ей дополнительные преимущества в утилизации питательных веществ окружающей среды.
Пили 2 типа (половые, F-пили, конъюгативные – sex pili) участвуют в конъюгации бактерий, обеспечивающей перенос части генетического материала от донорской клетке к реципиентной. Они имеются только у бактерий-доноров в ограниченном количестве (1-4 на клетку), более длинные (0,5-10 мкм). Отличительной особенностью половых пилей является взаимодействие с особыми «мужскими» сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Выявить пили можно при электронной микроскопии.
Жгутики и пили бактерий. Электронная микроскопия. (Атлас по медицинской микробиологии, вирусологии и иммунологии / Под ред. А.А. Воробьева, А.С. Быкова – М.: Медицинское информационное агентство, 2003.-236 с.).Escherichia coli. Электронная микроскопия. Адсорбция фага ms2 на f-пили. х100000.«Авакян А.А., Кац Л.Н., Павлова И.Б. Атлас анатомии бактерий, патогенных для человека и животных. М «Медицина».-1972.-183 с.»
функции и методы их выявления
Наряду со жгутиками прокариоты могут обладать и другими внеклеточными образованиями. В середине ХХ века было установлено, что бактерии способны формировать специфическую группу поверхностных образований. Их называли ворсинками, ресничками, фимбриями. Сегодня их называют пилями бактерий.

Внешний вид фимбрий
Внешне пили, или фимбрии, как их называли до 1956 г., выглядят как микроскопические волоски, покрывающие клетку бактерии. На 1 клетку прокариота может приходиться от нескольких единиц до тысяч ворсинок.
Хотя они, как и жгутики, являются поверхностными образованиями, однако между собой имеют больше различий, чем сходств.
По размеру пили намного меньше жгутиков, в среднем в 3 раза тоньше (не более 10 нм), и длиной не превышают 1,5 мкм.
По строению, несмотря на то, что как пили, так и жгутики состоят из белковых клеток, они также различаются:
- пили, или фимбрии, представляют собой легкую цепочку проводящих белков цилиндрической формы, отходящую от поверхностного слоя клетки;
- жгутики являются более громоздкими по строению, с наличием сложных структур (стержень, базальное тело, кольца и другое).
 Столь явное различие в строении поверхностных образований прокариотов связано с совершенно разными задачами, которые они решают в процессе жизнедеятельности бактериальной клетки.
Столь явное различие в строении поверхностных образований прокариотов связано с совершенно разными задачами, которые они решают в процессе жизнедеятельности бактериальной клетки.
Для чего прокариотам пили
К примеру, если жгутики бактерий обеспечивают возможность передвигаться, то фимбрии не имеют никакого отношения к перемещению в пространстве и присутствуют как у движущихся, так и у неподвижных бактерий.
Неподтвержденным исключением можно считать участие пилей 4 типа в скользящем движении прокариотов всей колонией, характерном для многих протеобактерий (и миксобактерий), скользящих бактерий, циано-, несерных бактерий и других.
В отличие от жгутиков функции пилей бактерий изучены достаточно слабо, но совершенно очевидно, что одной из них является способность обеспечить прикрепление клетки бактерии к питательному субстрату.
Разные типы ворсинок
Пили не являются однородными образованиями, их различают как минимум 4 типа, каждый из которых выполняет свои функции, причем одна клетка может являться носителем несколько разных типов фимбрий.
Пили 1 типа
Фимбрии бактерий 1 типа образуются из пилина (белок) и отличаются крайне прочной связью с прокариотом. Чтобы отделить такую фимбрию от бактериальной клетки, требуются усилия, многократно превышающие необходимое воздействие для отделения половых пилей или жгутиков.

Для пилей 1 типа характерно расположение перитрихиально – по всей поверхности бактерии.
Исследования методами выявления свойств показали, что пили 1 типа являются химически устойчивыми образованиями – они инертны к растворам щелочей, мочевине и трипсину (фермент, расщепляющий белки).
Разрушаются пили 1 типа при кипячении в растворах с высокой кислотностью, при этом методе воздействия происходит необратимое разрушение (денатурация) белка, образующего фимбрию.
Характерной особенностью пилей 1 типа является:
- способность образовывать пленки и придавать бактериям гидрофобные свойства;
- способность вызывать агглютинацию эритроцитов (выпадение в осадок в результате склеивания) под действием агглютининов.

Органоиды бактерий
Основными функциями являются:
- адгезивная – прикрепление бактерий к субстратам;
- защитная – объединение клеток прокариотов, получивших гидрофобные свойства, в группы;
- участие в процессах метаболизма клетки – увеличение всасывающей поверхности.
Ворсинки 2 типа
Эта группа имеет очень много общего с предыдущей, однако не обладает характерными особенностями 1 типа – пили не участвуют в формировании пленок и не приклеиваются к эритроцитам (агглютинация), провоцируя выпадение их в осадок.
Столь близкое сходство позволяет предположить, что пили 2 типа являются мутантной формой 1 типа.
Половые фимбрии (3 тип)
Современные методы выявления позволили определить, что при горизонтальном переносе генетического материала (конъюгации) ключевую роль играют половые пили.
Возможность непосредственного контакта двух бактериальных клеток с последующей конъюгацией была выявлена в 50-х годах прошлого века в результате исследований двух американских биохимиков – Д.Ледербейга и Э.Тейтема. Данный процесс имеет большое практическое значение, так как позволяет производить обмен наследственными признаками организмам, размножающимся только прямым делением.
Половые фимбрии, их называют F-пили, присутствуют только у бактериальных штаммов, обладающих фактором трансмиссивности – это может быть автономный репликон или его часть.
F-пили представляют собой цилиндрические белковые образования большего диаметра, чем пили 1 или 2 типа, расположенные перпендикулярно к поверхности.
 Формирование пиля осуществляется на поверхности цитоплазматической мембраны в точках контакта ее с вешней оболочкой. Сформированная трубочка проходит сквозь слои муреина и внешнюю мембрану.
Формирование пиля осуществляется на поверхности цитоплазматической мембраны в точках контакта ее с вешней оболочкой. Сформированная трубочка проходит сквозь слои муреина и внешнюю мембрану.
В случае потери F-пили восстанавливаются – в течение 30 секунд пиль достигает половины своей величины. Для формирования полноценной трубочки необходимо от 4 до 5 минут.
Современные методы выявления позволили определить, что на поверхности бактерии F-пили сохраняются в течение 5 минут, после чего сбрасываются, и процесс повторяется.
F-пили значительно отличаются от ворсинок 1 и 2 типа как по строению, так и по свойствам.
В отличие от последних F-пили легко отделяются от бактериальной клетки даже при незначительном встряхивании.
Методами физико-химического анализа было определено, что в составе F-пили отсутствует целый ряд α-аминокислот, характерных для белка пилей 1 типа, но через ковалентную связь присоединены остаток D-глюкозы и две фосфатные группы.
В связи с иной химической структурой на F-пили не адсорбируются обычные фаги, а только специфичные для них, называемые мужскими фагами.
Участие F-пили в процессе передачи информации
Процесс передачи части генетической информации подразумевает наличие пары клетка-донор и клетка-реципиент.
- Первоначально клетка-донор формирует F-пиль.
- F-пиль донора фиксируется на клетке-реципиенте.
- В F-плазмиде донорной клетки осуществляется разрыв одной нити ДНК, которая передается реципиенту.
- Обе бактерии достраивают вторую цепочку ДНК и восстанавливают F- плазмиду. Клетка-реципиент превращается в донора.

Микроскопические методы исследований позволили определить, что образование F-пилей характерно только для растущих и активных клеток, при переходе в стационарную фазу роста бактерии теряют свою способность образовывать половые пили и становятся плохими донорами.
Специфическая направленность фимбрий 4 типа
Пили 4 типа принимают участие в обеспечении разновидности скользящего движения бактерий всей колонией.
 Сам процесс скольжения с участием пилей 4 группы предполагает наличие 2 систем движения:
Сам процесс скольжения с участием пилей 4 группы предполагает наличие 2 систем движения:
- А-система — секретирует слизь на полюсе направления движения микроорганизма.
- S-система – роение; обеспечивается последовательным сокращением и удлинением пилей 4 типа, подобное подтягиванию.
Механизм данного вида бактериального движения на сегодняшний день находится в процессе изучения, и большинство выводов носят предположительный характер.

Образование высшее филологическое. В копирайтинге с 2012 г., также занимаюсь редактированием/размещением статей. Увлечения — психология и кулинария.
Пили — это… Что такое Пили?
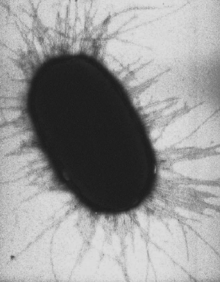 E. coli с перитрихиально расположенными многочисленными пилями
E. coli с перитрихиально расположенными многочисленными пилямиПили, фимбрии или ворсинки — поверхностные структуры, присутствующие у многих бактериальных клеток и представляющие собой прямые белковые цилиндры длиной 1—1,5 мкм и диаметром 7—10 нм. Различаются по строению и назначению, причём у одной бактерии могут присутствовать несколько их типов. Во многих случаях функции пилей не до конца установлены, но всегда они так или иначе участвуют в прикреплении бактериальной клетки к субстрату.
Наибольшее количество сведений о данных структурах собрано для пилей кишечных бактерий, прежде всего Escherichia coli.
Пили типа 1
Пили типа 1 прочно связаны с клеткой, и для того, чтобы отсоединить их от неё, нужны значительные усилия, большие, нежели для удаления жгутиков или половых пилей. Пили данного типа также устойчивы и к химическим воздействиям — сохраняются в 6 М мочевине, 1 N NаОН, устойчивы к додецилсульфату натрия и трипсину. Эти пили разрушаются только при кипячении в растворе с низким значением pH, что вызывает необратимую денатурацию белка. Белок, образующий пили общего типа 1, имеет молекулярную массу 17 кДа.
Пили типа 1 располагаются перитрихиально, то есть по всей поверхности бактерии. У одной клетки может быть 50—400 пилей длиной до 1,5 мкм. Диаметр этих пилей около 7 нм, а отверстия — 2,0—2,5 нм.
Формирование пилей общего типа 1 определяется генами, расположенными в хромосоме. Их активность подвержена фазовым вариациям, то есть ген может быть активен либо нет. Обычно в культуре присутствуют как клетки, имеющие много пилей общего типа 1, так и лишенные их. Клетки, находящиеся в той или иной фазе, могут быть легко выведены. Размножению клеток, лишенных пилей, способствует выращивание культуры на агаре, тогда как клетки с пилями получают преимущество при выращивании культуры в жидкой среде без аэрации. При этом они образуют пленку. Пили типа 1 придают бактериям гидрофобность, снижают их электрофоретическую подвижность. Они вызывают агглютинацию эритроцитов за счет того, что такие бактерии приклеиваются к эритроцитам (так же, как к другим клеткам животных), а также к клеткам растений и грибов, к неорганическим частицам. В присутствии маннозы нарушается гемагглютинация и прикрепление бактерий к животным клеткам вообще, поскольку пили типа 1 прикрепляются к поверхностным рецепторам, содержащим маннозу. В присутствии маннозы соответствующие участки пилей заняты её молекулами. Адгезивность пилей зависит также от гидрофобности образующего их белка пилина. С маннозными рецепторами реагируют участки пилей, расположенные по всей их поверхности, тогда как за гидрофобные взаимодействия ответственны окончания пилей.
Пили типа 2
Пили типа 2 сходны с пилями 1-го типа, но не вызывают агглютинации эритроцитов, не способствуют образованию бактериями пленки в жидкой среде. Антигенно они близки к пилям 1-го типа и, по-видимому, представляют собой их мутантную форму. Описан и еще ряд вариантов пилей, близких к пилям 1-го типа. Связи пилей общего типа 1 с патогенностью у штаммов Е. coli не удается обнаружить. У энтеропатогенных штаммов обычно образуются другие пили, кодируемые плазмидными генами. Известно несколько типов таких пилей, причем обнаруживается связь типа пилей со специфичностью бактерий в отношении тех или иных животных.
Другие типы пилей
Пили, известные как антигены К88 и К99, тоньше и лабильнее пилей 1-го типа. Они вызывают гемагглютинацию, устойчивую к маннозе, и способствуют прикреплению бактерий к клеткам кишечного эпителия животных, но не человека. Пили 987Р определяют способность Е. соli прикрепляться к эпителию тонкого кишечника новорожденных свиней; морфологически они похожи на пили 1-го типа. Пили, определяемые генетическим фактором СFА/1, вызывают агглютинацию человеческих эритроцитов и найдены у патогенных для человека штаммов. Молекулярная масса белков пилинов, кодируемых плазмидными генами, 14,5—26,2 кДа. У энтеропатогенных штаммов Е. соli пили являются одним из факторов патогенности, обеспечивающим им возможность прикрепления к клеткам кишечного эпителия. Колонизация бактериями эпителия способствует эффективному взаимодействию выделяемого ими энтеротоксина с клетками эпителия. В результате происходит нарушение водного обмена ткани, что клинически проявляется как диарея. При этом бактерии энергично размножаются в тонком кишечнике, а затем в большом количестве выносятся в окружающую среду, что способствует их распространению.
Половые пили
Половые пили Е. соli образуются у клеток донорских штаммов, отличающихся от изогенных реципиентных наличием у клеток особого генетического детерминанта — полового фактора, или фактора трансмиссивности, который либо является автономным репликоном (F-фактор), либо входит в состав автономного репликона, либо интегрирован с бактериальной хромосомой. Фактор трансмиссивности находится в составе плазмид — факторов множественной устойчивости к антибиотикам (R-факторы), факторов колициногенности и ряда других плазмид. Половые пили отличаются от пилей общего типа по строению и антигенной специфичности, пили, кодируемые различными генетическими детерминантами, также различны.
Половые F-пили, определяемые F-факторами, представляют собой белковые цилиндры, перпендикулярные поверхности клетки, толщиной 8,5—9,5 нм и длиной до 1,1 мкм. Они легко могут быть отделены от клетки при встряхивании бактериальной массы. F—пили образованы белком с молекулярной массой 11,8 кДа. В составе F—пилина отсутствуют пролин, цистеин, гистидин, аргинин. К молекуле пилина присоединены две фосфатные группы и остаток D-глюкозы, связанные с белком ковалентными связями. Пилин содержит довольно много кислых и гидрофобных аминокислот. Он синтезируется на рибосомах, связанных с цитоплазматической мембраной и в цитоплазме не обнаруживается. Пул пилина, видимо, накапливается в цитоплазматической мембране. Его молекулы в процессе синтеза содержат дополнительную сигнальную последовательность аминокислот, отщепляющуюся при транспорте через мембрану. F—пили легко диссоциируют в растворах додецилсульфата натрия и разрушаются органическими растворителями, что связано с гидрофобностью пилина. Бактерии, имеющие F—пили, приобретают новый антиген, у них изменяется поверхностный заряд. Бактерии с F-пилями малоподвижны, проявляют тенденцию к автоагглютинации, например, при понижении значения рН среды. Это также происходит за счет богатства пилина кислыми и гидрофобными аминокислотами. F—фактор интересен еще и потому, что иногда (примерно в 1 случае из 100000) он встраивается в молекулу основной ДНК клетки-хозяина. Тогда при конъюгации переносится не только F—фактор, но, также и остальная ДНК. Этот процесс занимает примерно 90 минут, но клетки могут расходиться и раньше, до полного обмена ДНК. Такие штаммы постоянно передают всю или большую часть своей ДНК другим клеткам. Эти штаммы называются Hrf-штаммами (High frequency recombination), потому что донорная ДНК таких штаммов рекомбинирует с ДНК реципиента.
Для образования F-пилей необходима активность, по крайней мере, 13 генов. Сборка трубочек пилей происходит на цитоплазматической мембране в местах ее контакта с внешней мембраной. Трубочка пили проходит через слои муреина и внешнюю мембрану. Для сборки и сохранения пилей необходима энергия. Образованию пилей препятствуют цианид, динитрофенол, азид натрия. Возможно, в процессе сборки происходит фосфорилирование пилина. Обычно клетки с дерепрессированным F—фактором образуют 1—2 пили, а в анаэробных условиях и на богатой среде — до 5 пилей. Причина стимуляции пилеобразования в анаэробных условиях неизвестна. У клеток с оторванными пилями быстро отрастают новые, за 30 секунд пиля достигает 1/2 нормальной длины, а полностью формируется за 4—5 мин. Сформированные пили сохраняются на поверхности клетки 4—5 мин, а затем сбрасываются. Это свидетельствует в пользу точки зрения о том, пили — активные образования. Пили, определяемые фактором Соl I, образованы иным пилином, на них не адсорбируются фаги, специфичные для F—пилей, но имеются специфичные для них фаги. Так называемые мужские фаги адсорбируются на половых пилях, РНК-содержащие фаги — на их боковых поверхностях и нитчатые фаги, содержащие одноцепочечную ДНК, — на кончиках этих пилей. Нитчатый фаг препятствует конъюгации.
При конъюгации к реципиентной клетке присоединяется конец половой пили, при этом рецептором служит белок внешней мембраны реципиентной клетки. Сначала этот контакт не очень прочный и легко может быть нарушен при гидродинамических воздействиях. При этом пары распадаются при множественном заражении РНК-содержащими фагами или в присутствии ионов Zn2+. Через несколько минут контакт становится более прочным, происходит сближение клеток и образование между ними цитоплазматического мостика. Имеются данные, свидетельствующие о том, что передача ДНК может происходить и без образования цитоплазматического мостика, а непосредственно через отверстие в пиле. Инактивация пилей антисывороткой и любые повреждающие их воздействия приводят к нарушению процесса конъюгации, в то время как нарушение целостности внешней мембраны или муреинового слоя до некоторого предела влияют на донорские свойства клетки, имеющей пили. После установления контакта с реципиентной клеткой черв пилю в донорскую клетку передается сигнал, вызывающий начало конъюгационного синтеза ДНК. Механизм работы половых пилей еще окончательно не установлен. Ряд наблюдений свидетельствует в пользу модели, предполагающей активную функцию пилей. Согласно этой точке зрения после установления контакта с клеткой реципиента или с вирусом пиля сокращается или втягивается в клетку. Эта модель подтверждается как косвенными, так и прямыми наблюдениями. На электронно-микроскопических препаратах можно проследить, как после адсорбции нитчатого мужского фага на их кончиках пили укорачиваются, а затем нити фага оказываются на поверхности клетки. Сокращение пилей вызыват KCN или арсенат. После воздействия этими ингибиторами пили не обнаруживаются ни на поверхности клеток, ни в окружающей среде, но можно наблюдать адсорбцию на поверхности клеток мужских фагов и антител, специфичных к концам пилей, то есть их кончики, видимо, продолжают выступать над поверхностью клетки. При фаговой инфекции в дальнейшем происходит растворение белковой оболочки нитчатого фага в цитоплазматической мембране бактерии и освобождение его ДНК в цитоплазму. При инфицировании РНК-содержащими мужскими фагами сначала образуется комплекс фаговой РНК с пилином, а фаговый капсид освобождается в среду.
Обычно синтез пилина находится под контролем цитоплазматических репрессоров. В некоторых случаях удается наблюдать определенные закономерности в регуляции образования пилей. Так, в случае Соl I—фактора каждая клетка, получившая при конъюгации плазмиду Соl I, образует пили, их активное образование происходит у клеток 4—8 последующих генераций. Однако затем только единичные клетки в популяции образуют пили, поскольку у большинства бактерий синтез пилина репрессирован. Подобная репрессия, как считают, имеет приспособительное значение, поскольку клетки без пилей не чувствительны к мужским бактериофагам, которые могли бы уничтожить всю популяцию. Единичные клетки с пилями способны обеспечить конъюгацию. При контакте таких клеток с популяциями реципиентных бактерий начинается лавинообразное распространение плазмиды, поскольку образование пилей сначала не репрессировано.
Половые пили обычно образуют только активно растущие клетки, клетки из культуры, находящейся в стационарной фазе роста, обычно лишены пилей и являются плохими донорами.
Как уже было отмечено, существует много более или менее различающихся плазмид, способных определять образование половых пилей, которые также несколько различаются. Рецепторы на поверхности реципиентных клеток обладают разной степенью сродства к разным пилям, что может сильно влиять на эффективность конъюгации бактерий.
Пили, подобные пилям E. coli, образуют и другие представители Enterobacteriaceae. Половые пили имеют Vibrio, Pasteurella, Aeromonas, Pseudomonas.
Литература
- Громов Б. В. Строение Бактерий: Учеб пособие. — Л.: Изд-во Ленингр. ун-та, 1985. — 192 с.
- Н. Грин, У. Стаут, Д. Тейлор Биология: В 3-х т. Т. 1: М.: Мир, 1996. — 368 с.
Ворсинки (фимбрии, пили) бактерий
Ворсинки (фимбрии, пили) бактерий
К поверхностным структурам бактериальной клетки относятся также ворсинки (фимбрии, пили) ( рис. 4 , 6). Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм. Ворсинки построены из одного вида белка — пилина — и представляют собой прямые белковые цилиндры, отходящие от поверхности клетки. Они, как правило, тоньше жгутиков (диаметр — 5-10 нм, длина 0,2-2,0 мкм), расположены перитрихиально или полярно. Больше всего сведений имеется о ворсинках Е. coli . У этой бактерии описаны ворсинки общего типа и половые.
Ворсинки общего типа придают бактериям свойство гидрофобности, обеспечивают их прикрепление к клеткам растений, грибов и неорганическим частицам, принимают участие в транспорте метаболитов. Через ворсинки в клетку могут проникать вирусы .
Наиболее хорошо изучены половые ворсинки, или F-пили, принимающие участие в половом процессе бактерий. F-пили необходимы клетке-донору для обеспечения контакта между ней и реципиентом и в качестве конъюгационного тоннеля, по которому происходит передача ДНК. Ворсинки нельзя считать обязательной клеточной структурой, так как и без них бактерии хорошо растут и размножаются.
Фимбрии (пили) — нитевидные белковые органеллы, покрывающих всю поверхность бактериальной клетки — антигены фактора колонизации . Эти тонкие структуры позволяют бактерии прикрепляться к эпителиальным клеткам и препятствуют ее захвату нейтрофилами
Фимбрии состоят из множества одинаковых белковых субъединиц. Эта субъединица называется пилином (молекулярная масса 17000-30000). В составе пилина есть консервативные и вариабельные участки. Перестройки хромосом, ведущие к экспрессии любого из множества неактивных генов пилина, сопровождаются изменениями антигенного состава фимбрий.
При электронной микроскопии фимбрии выглядят как похожие на волоски выросты, проникающие через наружную мембрану. Они могут располагаться на одном конце клетки либо более равномерно по всей ее поверхности. У отдельной клетки может быть несколько сотен фимбрий, которые выполняют различные функции.
У некоторых фимбрий (например, у дигалактозидсвязывающих фимбрий Escherichia coli ) на апикальном конце находятся специальные белки, играющие важную роль во взаимодействии с рецепторами клеток.
Считается, что главная функция фимбрий — обеспечение фиксации бактерий в тканях. Адгезия микробная: специфичность тканевая и видоваяАдгезия микробная: специфичность тканевая и видовая
Ссылки:
Все ссылкиПили — Википедия. Что такое Пили
E. coli с перитрихиально расположенными многочисленными пилямиПили (пилус), фимбрии или ворсинки — поверхностные структуры, присутствующие у многих бактериальных клеток и представляющие собой прямые белковые цилиндры длиной 1—1,5 мкм и диаметром 7—10 нм. Различаются по строению и назначению, причём у одной бактерии могут присутствовать несколько их типов. Во многих случаях функции пилей не до конца установлены, но всегда они так или иначе участвуют в прикреплении бактериальной клетки к субстрату.
Наибольшее количество сведений о данных структурах собрано для пилей кишечных бактерий, прежде всего Escherichia coli.
Пили типа 1
Пили типа 1 прочно связаны с клеткой, и для того, чтобы отсоединить их от неё, нужны значительные усилия, большие, нежели для удаления жгутиков или половых пилей. Пили данного типа также устойчивы и к химическим воздействиям — сохраняются в 6 М мочевине, 1 М NаОН, устойчивы к додецилсульфату натрия и трипсину. Эти пили разрушаются только при кипячении в растворе с низким значением pH, что вызывает необратимую денатурацию белка. Белок, образующий пили общего типа 1, имеет молекулярную массу 17 кДа.
Пили типа 1 располагаются перитрихиально, то есть по всей поверхности бактерии. У одной клетки может быть 50—400 пилей длиной до 1,5 мкм. Диаметр этих пилей около 7 нм, а отверстия — 2,0—2,5 нм.
Формирование пилей общего типа 1 определяется генами, расположенными в хромосоме. Их активность подвержена фазовым вариациям, то есть ген может быть активен либо нет. Обычно в культуре присутствуют как клетки, имеющие много пилей общего типа 1, так и лишенные их. Клетки, находящиеся в той или иной фазе, могут быть легко выведены. Размножению клеток, лишенных пилей, способствует выращивание культуры на агаре, тогда как клетки с пилями получают преимущество при выращивании культуры в жидкой среде без аэрации. При этом они образуют пленку. Пили типа 1 придают бактериям гидрофобность, снижают их электрофоретическую подвижность. Они вызывают агглютинацию эритроцитов за счет того, что такие бактерии приклеиваются к эритроцитам (так же, как к другим клеткам животных), а также к клеткам растений и грибов, к неорганическим частицам. В присутствии маннозы нарушается гемагглютинация и прикрепление бактерий к животным клеткам вообще, поскольку пили типа 1 прикрепляются к поверхностным рецепторам, содержащим маннозу. В присутствии маннозы соответствующие участки пилей заняты её молекулами. Адгезивность пилей зависит также от гидрофобности образующего их белка пилина. С маннозными рецепторами реагируют участки пилей, расположенные по всей их поверхности, тогда как за гидрофобные взаимодействия ответственны окончания пилей.
Пили типа 2
Пили типа 2 сходны с пилями 1-го типа, но не вызывают агглютинации эритроцитов, не способствуют образованию бактериями пленки в жидкой среде. Антигенно они близки к пилям 1-го типа и, по-видимому, представляют собой их мутантную форму. Описан и еще ряд вариантов пилей, близких к пилям 1-го типа. Связи пилей общего типа 1 с патогенностью у штаммов Е. coli не удается обнаружить. У энтеропатогенных штаммов обычно образуются другие пили, кодируемые плазмидными генами. Известно несколько типов таких пилей, причем обнаруживается связь типа пилей со специфичностью бактерий в отношении тех или иных животных.
Другие типы пилей
Пили, известные как антигены К88 и К99, тоньше и лабильнее пилей 1-го типа. Они вызывают гемагглютинацию, устойчивую к маннозе, и способствуют прикреплению бактерий к клеткам кишечного эпителия животных, но не человека. Пили 987Р определяют способность Е. соli прикрепляться к эпителию тонкого кишечника новорожденных свиней; морфологически они похожи на пили 1-го типа. Пили, определяемые генетическим фактором СFА/1, вызывают агглютинацию человеческих эритроцитов и найдены у патогенных для человека штаммов. Молекулярная масса белков пилинов, кодируемых плазмидными генами, 14,5—26,2 кДа. У энтеропатогенных штаммов Е. соli пили являются одним из факторов патогенности, обеспечивающим им возможность прикрепления к клеткам кишечного эпителия. Колонизация бактериями эпителия способствует эффективному взаимодействию выделяемого ими энтеротоксина с клетками эпителия. В результате происходит нарушение водного обмена ткани, что клинически проявляется как диарея. При этом бактерии энергично размножаются в тонком кишечнике, а затем в большом количестве выносятся в окружающую среду, что способствует их распространению.
Половые пили
Половые пили Е. соli образуются у клеток донорских штаммов, отличающихся от изогенных реципиентных наличием у клеток особого генетического детерминанта — полового фактора, или фактора трансмиссивности, который либо является автономным репликоном (F-фактор), либо входит в состав автономного репликона, либо интегрирован с бактериальной хромосомой. Фактор трансмиссивности находится в составе плазмид — факторов множественной устойчивости к антибиотикам (R-факторы), факторов колициногенности и ряда других плазмид. Половые пили отличаются от пилей общего типа по строению и антигенной специфичности, пили, кодируемые различными генетическими детерминантами, также различны.
Половые F-пили, определяемые F-факторами, представляют собой белковые цилиндры, перпендикулярные поверхности клетки, толщиной 8,5—9,5 нм и длиной до 1,1 мкм. Они легко могут быть отделены от клетки при встряхивании бактериальной массы. F—пили образованы белком с молекулярной массой 11,8 кДа. В составе F—пилина отсутствуют пролин, цистеин, гистидин, аргинин. К молекуле пилина присоединены две фосфатные группы и остаток D-глюкозы, связанные с белком ковалентными связями. Пилин содержит довольно много кислых и гидрофобных аминокислот. Он синтезируется на рибосомах, связанных с цитоплазматической мембраной и в цитоплазме не обнаруживается. Пул пилина, видимо, накапливается в цитоплазматической мембране. Его молекулы в процессе синтеза содержат дополнительную сигнальную последовательность аминокислот, отщепляющуюся при транспорте через мембрану. F—пили легко диссоциируют в растворах додецилсульфата натрия и разрушаются органическими растворителями, что связано с гидрофобностью пилина. Бактерии, имеющие F—пили, приобретают новый антиген, у них изменяется поверхностный заряд. Бактерии с F-пилями малоподвижны, проявляют тенденцию к автоагглютинации, например, при понижении значения рН среды. Это также происходит за счет богатства пилина кислыми и гидрофобными аминокислотами. F—фактор интересен еще и потому, что иногда (примерно в 1 случае из 100000) он встраивается в молекулу основной ДНК клетки-хозяина. Тогда при конъюгации переносится не только F—фактор, но, также и остальная ДНК. Этот процесс занимает примерно 90 минут, но клетки могут расходиться и раньше, до полного обмена ДНК. Такие штаммы постоянно передают всю или большую часть своей ДНК другим клеткам. Эти штаммы называются Hrf-штаммами (High frequency recombination), потому что донорная ДНК таких штаммов рекомбинирует с ДНК реципиента.
Для образования F-пилей необходима активность, по крайней мере, 13 генов. Сборка трубочек пилей происходит на цитоплазматической мембране в местах её контакта с внешней мембраной. Трубочка пили проходит через слои муреина и внешнюю мембрану. Для сборки и сохранения пилей необходима энергия. Образованию пилей препятствуют цианид, динитрофенол, азид натрия. Возможно, в процессе сборки происходит фосфорилирование пилина. Обычно клетки с дерепрессированным F—фактором образуют 1—2 пили, а в анаэробных условиях и на богатой среде — до 5 пилей. Причина стимуляции пилеобразования в анаэробных условиях неизвестна. У клеток с оторванными пилями быстро отрастают новые, за 30 секунд пиля достигает 1/2 нормальной длины, а полностью формируется за 4—5 мин. Сформированные пили сохраняются на поверхности клетки 4—5 мин, а затем сбрасываются. Это свидетельствует в пользу точки зрения о том, пили — активные образования. Пили, определяемые фактором Соl I, образованы иным пилином, на них не адсорбируются фаги, специфичные для F—пилей, но имеются специфичные для них фаги. Так называемые мужские фаги адсорбируются на половых пилях, РНК-содержащие фаги — на их боковых поверхностях и нитчатые фаги, содержащие одноцепочечную ДНК, — на кончиках этих пилей. Нитчатый фаг препятствует конъюгации.
При конъюгации к реципиентной клетке присоединяется конец половой пили, при этом рецептором служит белок внешней мембраны реципиентной клетки. Сначала этот контакт не очень прочный и легко может быть нарушен при гидродинамических воздействиях. При этом пары распадаются при множественном заражении РНК-содержащими фагами или в присутствии ионов Zn2+. Через несколько минут контакт становится более прочным, происходит сближение клеток и образование между ними цитоплазматического мостика. Имеются данные, свидетельствующие о том, что передача ДНК может происходить и без образования цитоплазматического мостика, а непосредственно через отверстие в пиле. Инактивация пилей антисывороткой и любые повреждающие их воздействия приводят к нарушению процесса конъюгации, в то время как нарушение целостности внешней мембраны или муреинового слоя до некоторого предела влияют на донорские свойства клетки, имеющей пили. После установления контакта с реципиентной клеткой черв пилю в донорскую клетку передается сигнал, вызывающий начало конъюгационного синтеза ДНК. Механизм работы половых пилей еще окончательно не установлен. Ряд наблюдений свидетельствует в пользу модели, предполагающей активную функцию пилей. Согласно этой точке зрения после установления контакта с клеткой реципиента или с вирусом пиля сокращается или втягивается в клетку. Эта модель подтверждается как косвенными, так и прямыми наблюдениями. На электронно-микроскопических препаратах можно проследить, как после адсорбции нитчатого мужского фага на их кончиках пили укорачиваются, а затем нити фага оказываются на поверхности клетки. Сокращение пилей вызыват KCN или арсенат. После воздействия этими ингибиторами пили не обнаруживаются ни на поверхности клеток, ни в окружающей среде, но можно наблюдать адсорбцию на поверхности клеток мужских фагов и антител, специфичных к концам пилей, то есть их кончики, видимо, продолжают выступать над поверхностью клетки. При фаговой инфекции в дальнейшем происходит растворение белковой оболочки нитчатого фага в цитоплазматической мембране бактерии и освобождение его ДНК в цитоплазму. При инфицировании РНК-содержащими мужскими фагами сначала образуется комплекс фаговой РНК с пилином, а фаговый капсид освобождается в среду.
Обычно синтез пилина находится под контролем цитоплазматических репрессоров. В некоторых случаях удается наблюдать определенные закономерности в регуляции образования пилей. Так, в случае Соl I—фактора каждая клетка, получившая при конъюгации плазмиду Соl I, образует пили, их активное образование происходит у клеток 4—8 последующих генераций. Однако затем только единичные клетки в популяции образуют пили, поскольку у большинства бактерий синтез пилина репрессирован. Подобная репрессия, как считают, имеет приспособительное значение, поскольку клетки без пилей не чувствительны к мужским бактериофагам, которые могли бы уничтожить всю популяцию. Единичные клетки с пилями способны обеспечить конъюгацию. При контакте таких клеток с популяциями реципиентных бактерий начинается лавинообразное распространение плазмиды, поскольку образование пилей сначала не репрессировано.
Половые пили обычно образуют только активно растущие клетки, клетки из культуры, находящейся в стационарной фазе роста, обычно лишены пилей и являются плохими донорами.
Как уже было отмечено, существует много более или менее различающихся плазмид, способных определять образование половых пилей, которые также несколько различаются. Рецепторы на поверхности реципиентных клеток обладают разной степенью сродства к разным пилям, что может сильно влиять на эффективность конъюгации бактерий.
Пили, подобные пилям E. coli, образуют и другие представители Enterobacteriaceae. Половые пили имеют Vibrio, Pasteurella, Aeromonas, Pseudomonas.
Литература
- Громов Б. В. Строение Бактерий: Учеб пособие. — Л.: Изд-во Ленингр. ун-та, 1985. — 192 с.
- Н. Грин, У. Стаут, Д. Тейлор Биология: В 3-х т. Т. 1: М.: Мир, 1996. — 368 с.
Ссылки
Строение бактериальной клетки — Википедия
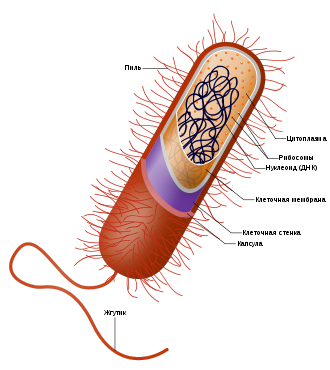 Схема строения бактериальной клетки
Схема строения бактериальной клеткиБактериа́льная кле́тка обычно устроена наиболее просто по сравнению с клетками других живых организмов. Бактериальные клетки часто окружает капсула, которая служит защитой от внешней среды. Для многих свободноживущих бактерий характерно наличие жгутиков для передвижения, а также ворсинок.
Для выведения веществ, в том числе факторов патогенности, в окружающую среду используются системы секреции. Клеточная стенка бактерий обычно содержит пептидогликан. По химическому составу клеточные мембраны бактерий гораздо разнообразнее мембран эукариотических клеток. В отличие от эукариот, бактерии не имеют ограниченного оболочкой ядра и, в большинстве случаев, каких-либо мембранных органелл. Вместе с тем у ряда бактерий имеются клеточные структуры, не имеющие аналогов в двух других доменах.
Геном бактерий состоит из суперскрученных кольцевых хромосом, связанных с гистонподобными белками, и меньших по размерам молекул ДНК — плазмид. Элементы цитоскелета играют важные роли в делении клеток, защите, поддержании формы и определении полярности у различных прокариот. Бактериальные рибосомы меньше рибосом эукариотического типа, но имеют сходный план строения.
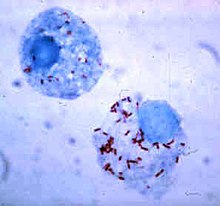 Риккетсии (красные точки) в клетках млекопитающего
Риккетсии (красные точки) в клетках млекопитающегоКак правило, размеры клеток бактерий находятся в пределах от 0,2 до 10 мкм. Существуют, однако, бактерии, видимые невооружённым глазом: клетки бактерии Epulopiscium fishelsoni, обитающей в кишечнике рыбы-хирурга, достигают до 600 мкм в длину и 100 мкм в диаметре, а клетки Thiomargarita namibiensis, населяющей прибрежные воды Намибии, достигает 400—750 мкм в диаметре[1].
Бактерий, клетки которых составляют менее 0,5 мкм в диаметре, называют нанобактериями, или ультрамикробактериями[en], они даже способны проходить через мембранные фильтры[en]. Среди ультрамикробактерий есть и свободноживущие виды, например, морская бактерия Sphingopyxis alaskensis, и непатогенные эндосимбионты, например, представители рода Holospora, размножающиеся в микро- или макронуклеусе инфузории-туфельки Paramecium caudatum. Многие ультрамикробактерии ведут паразитический образ жизни, в их числе микоплазмы, хламидии и риккетсии. Ультрамикробактерия Bdellovibrio bacteriovorus размножается в периплазматическом пространстве клеток других бактерий и питается продуктами лизиса хозяйской клетки, за что её часто относят к хищным бактериям[2].
 Различные морфотипы бактерий
Различные морфотипы бактерийФорма бактериальной клетки является диагностическим признаком и применяется в их классификации. Чаще всего бактериальные клетки имеют сферическую (кокки) или палочковидную (бациллы) формы, некоторые имеют форму, промежуточную между сферической и палочковидной, и называются коккобациллами. Многие бактерии имеют нитевидную или извитую форму — в виде запятой (вибрионы), спирали (спириллы[en]) или вытянутую, закрученную наподобие спирали ДНК[en] (спирохеты)[3].
Часто бактериальные клетки образуют устойчивые сочетания, такие как пары палочек (диплобациллы) или кокков (диплококки), цепочки палочек (стрептобациллы) или кокков (стрептококки), тетрады, пакеты из 4, 8 и более кокков (сарцины), гроздья (стафилококки). Некоторые бактерии образуют розетки, плоские таблички, сети, а также прямые или ветвящиеся трихомы — цепочки плотно примыкающих друг к другу клеток.
Известны бактерии с клетками весьма необычной формы (например, звёздчатые), некоторые бактерии (Corynebacterium[en]*, Mycobacterium, Nocardia[en]) меняют морфологию в течение жизненного цикла. Актинобактерии формируют мицелий, представители рода Hyphomicrobium образуют гифы с почками[4]. Клетки некоторых бактерий (например, Caulobacter) несут стебельки и прочие придатки[5].
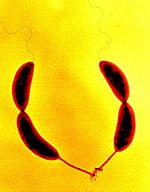 Две делящиеся клетки Caulobacter crescentus под микроскопом, виды стебельки на одном полюсе клеток и жгутики на противоположном
Две делящиеся клетки Caulobacter crescentus под микроскопом, виды стебельки на одном полюсе клеток и жгутики на противоположномВ отличие от многоклеточных организмов, у одноклеточных организмов (и бактерий в том числе) рост, то есть увеличение клетки в размерах, и размножение путём деления клеток тесно связаны[6]. Обычно бактериальные клетки делятся на две равноценные дочерние клетки. Сначала клетка удлиняется, в ней образуется поперечная перегородка. На завершающем этапе дочерние клетки расходятся. Отличительной чертой деления бактериальных клеток является непосредственное участие реплицированной ДНК в процессе деления[7]. В связи с тем, что в подавляющем большинстве случаев прокариотические клетки имеют клеточную стенку, бинарное деление сопровождается формированием септы — перегородки между дочерними клетками, которая затем расслаивается посередине. Процесс деления прокариотической клетки подробно изучен на примере E. coli[8].
В то же время есть примеры неравноценного деления. Например, у грамотрицательной бактерии Caulobacter crescentus[en] одна из дочерних клеток подвижная, у неё есть один жгутик для хемотаксиса. Вторая клетка остаётся прикреплённой к субстрату «стебельком». Подвижные клетки дифференцируются в клетки со стеблем после короткого периода свободного плавания. Репликация хромосом и деление клеток происходят только на стадии прикреплённой клетки[9].
В оптимальных условиях бактерии растут и делятся очень быстро, описан пример морской псевдомонады, популяция которой может удваиваться каждые 9,8 минуты[10].
 Структурная формула гопена — соединения группы гопаноидов
Структурная формула гопена — соединения группы гопаноидовКак любая живая клетка, бактериальная клетка окружена мембраной, которая представляет собой липидный бислой (её ещё называют цитоплазматической мембраной). Клеточная мембрана поддерживает осмотический баланс клетки, осуществляет разные виды транспорта, в том числе секрецию белков, задействована в образовании клеточной стенки и биосинтезе внеклеточных полимеров, а также получает регуляторные сигналы из внешней среды. Во многих случаях клеточная мембрана может участвовать в синтезе АТФ за счёт трансмембранного электрохимического градиента (протондвижущей силы). Мембрана бактериальной клетки участвует в репликации и разделении дочерних бактериальных хромосом при делении клетки, а также в передаче ДНК посредством трансдукции или конъюгации[11].
Помимо липидов, в состав бактериальных мембран входят различные белки. По химическому составу клеточные мембраны бактерий гораздо разнообразнее мембран эукариотических клеток. Мембранные липиды архей представлены ацил[en]— и алкилсодержащими глицеролипидами (в том числе фосфолипидами), а также полиизопреноидами. В отличие от эукариот, меняющих свойства липидного остова мембраны за счёт изменения соотношения между фосфолипидами и холестерином, бактерии изменяют свойства мембраны, варьируя жирные кислоты, входящие в состав липидов. Стероиды обнаруживаются в бактериальных мембранах чрезвычайно редко, и вместо стероидов мембраны содержат гопаноиды, представляющие собой пентациклические[en] углеводороды. Гопаноиды активно участвуют в регуляции физических свойств мембран бактериальных клеток[12].
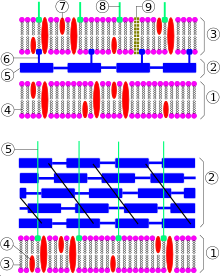 Схема строения клеточной стенки грамотрицательной (сверху) и грамположительной (снизу) бактерий. Сверху: 1 — клеточная мембрана, 2 — периплазматическое пространство, 3 — внешняя мембрана, 4 — фосфолипид, 5 — пептидогликан, 6 — липопротеин, 7 — белок, 8 — липополисахарид, 9 — порины. Снизу: 1 — клеточная мембрана, 2 — пептидогликан, 3 — фосфолипид, 4 — белок, 5 — липотейхоевая кислота
Схема строения клеточной стенки грамотрицательной (сверху) и грамположительной (снизу) бактерий. Сверху: 1 — клеточная мембрана, 2 — периплазматическое пространство, 3 — внешняя мембрана, 4 — фосфолипид, 5 — пептидогликан, 6 — липопротеин, 7 — белок, 8 — липополисахарид, 9 — порины. Снизу: 1 — клеточная мембрана, 2 — пептидогликан, 3 — фосфолипид, 4 — белок, 5 — липотейхоевая кислотаВ зависимости от типа строения клеточной стенки бактерии подразделяют на грамположительные и грамотрицательные (названия группам были даны вследствие их разного окрашивания по методу Грама). Большинство бактериальных клеток окружены жёсткой клеточной стенкой, состоящей из полимера пептидогликана, также известного как муреин. Пептидогликан состоит из полисахаридных цепей, скреплённых короткими пептидными сшивками. В большинстве случаев клеточная стенка жизненно необходима для бактерии, поэтому антибиотики, блокирующие её образование (формирование) (например, пенициллин), эффективны против самых разных бактерий. В старых культурах и при несбалансированном росте грамотрицательных бактерий появляются так называемые сферопласты — клетки, лишённые клеточной стенки или имеющие дефекты в ней. Однако, в отличие от протопластов, они взаимодействуют с бактериофагами, размножаются и при благоприятных условиях возвращаются в нормальное состояние. Сферопласты патогенных бактерий называются L-формами, которые получаются в лабораториях в отсутствие клеточной стенки только в изотонических растворах[13][14][15]. Некоторые бактерии, паразитирующие внутри эукариотических клеток, например, микоплазма, не имеют клеточной стенки[16].
Клеточная стенка механически стабилизирована и противостоит внутреннему давлению (тургорному давлению) бактериальной клетки, которое составляет от 2 до 25 атм[17]. Кроме того, она играет ключевую роль в поддержании формы бактериальной клетки[18]. Через слой пептидогликана, имеющий небольшие отверстия, могут проходить только относительно небольшие молекулы (массой до 50—60 кДа), причём размер проникающих молекул не зависит от толщины слоя пептидогликана. В связи с этим в тех случаях, когда через слой пептидогликана должны пройти большие молекулы, такие как белки жгутиков, пилей и ДНК при конъюгации, специфические гидролазы пептидогликана локально расширяют отверстия для их прохода[17].
Химическое строение[править | править код]
 Упрощённая схема строения пептидогликана
Упрощённая схема строения пептидогликанаГликановые цепи пептидогликана обычно состоят из повторяющегося дисахарида N-Ацетилглюкозамин-N-Ацетилмурамовой кислоты (NAG-NAM). В среднем нить образована 30 дисахаридами, но их количество может варьировать. К NAM присоединяется короткий пептид, состоящий из аминокислот D-глутаминовой кислоты, D-аланина и диаминопимелиновой кислоты (DAP) и синтезируемый вне рибосом. Аминогруппы DAP участвуют в образовании сшивок между полисахаридными цепями пептидогликана. При образовании сшивок последний D-аланин пептида высвобождается. У некоторых бактерий в пептидных мостиках присутствуют другие аминокислоты, а у грамположительных бактерий гликановые нити могут также соединяться с одним или несколькими остатками глицина в пептидных мостиках[19].
Грамположительные бактерии[править | править код]
 Окрашивание по Граму. Грамположительные кокки окрашены фиолетовым цветом, а грамотрицательные палочки — розовым
Окрашивание по Граму. Грамположительные кокки окрашены фиолетовым цветом, а грамотрицательные палочки — розовымУ грамположительных бактерий поверх мембраны есть (от 20 до 50 нм) оболочка из пептидогликана толщиной до 40 молекулярных слоёв[20]. Их положительная окраска по методу Грама связана с тем, что их толстая пептидогликановая клеточная стенка прочно связывает комплекс красителя генцианвиолета[en] с йодом, который не вымывается. Поэтому на препаратах грамположительные бактерии выглядят фиолетовыми (у грамотрицательных бактерий этот комплекс вымывается, и они приобретают цвет второго красителя, например сафранина)[21].
Кроме пептидогликана, в клеточной стенке грамположительных бактерий имеются тейхоевые кислоты, которые закрепляются на поверхности клетки, образуя связи с пептидогликаном. Липотейхоевые кислоты взаимодействуют с остатками жирных кислот клеточной мембраны. Тейхоевые и липотейхоевые кислоты представляют собой полианионы, состоящие из повторяющихся звеньев в виде фосфорилированных сахаров или остатков глицерина. Фосфатные группы в составе тейхоевых кислот могут быть заменены на глюкоуронат, в результате чего образуются тейхуроновые кислоты. Синтез тейхуроновых кислот запускается при фосфорном голодании. Блокировка синтеза тейхоевых кислот приводит к гибели бактерий, однако конкретные функции этих соединений точно не установлены[22]. Высказываются предположения, что они действуют наподобие пружин, делая возможным растяжение и сжатие клеточной стенки. Кроме того, за счёт своей полианионной природы тейхоевые кислоты прочно связывают ионы магния, поэтому могут выполнять в клетке роль ионообменника[23].
Поскольку у грамположительных бактерий слой пептидогликана не прикрыт сверху мембраной, перед ними стоит проблема удержания поверхностных белков. В ряде случаев поверхностные белки при помощи специальных ферментов пришиваются к фосфолипидам клеточной мембраны с образованием липопротеинов. Кроме того, поверхностные белки могут закрепляться на поверхности клетки за счёт связывания с пептидогликаном, которое обеспечивается ферментом сортазой[en]. Белки, предназначенные к связыванию с пептидогликаном, несут на N- и C-концах характерные последовательности, например, на N-конце находится сигнальный пептид, благодаря которому белок проходит через клеточную мембрану. Вблизи C-конца находится мотив, распознаваемый сортазой; в него вносится разрыв, после чего белок с отрезанным C-концевым фрагментом ковалентно пришивается к пептидогликану амидной связью[24].
У микобактерий, нокардий и коринебактерий 30 % вещества клеточной стенки составляют липиды, причём у некоторых микобактерий в ней также образованы воски. Такую обогащённую липидами клеточную стенку иногда называют микомембраной. Микомембрана защищает бактерий от неблагоприятных условий среды и антимикробных препаратов[25]. У бактерий родов Mycobacterium, Nocardia, Corynebacterium, Rhodococcus[en] и Caseobacter в клеточной стенке обнаруживаются миколовые кислоты. Помимо полисахаридов, у ряда патогенных грамположительных бактерий в клеточных стенках присутствуют белки, например, белок А у стафилококков, который служит важным антигеном. Кроме того, с клеточной стенкой временно, перед высвобождением в окружающую среду, связываются энтеротоксины[en][26].
Грамотрицательные бактерии[править | править код]
 Подробная схема строения клеточной стенки грамотрицательных бактерий
Подробная схема строения клеточной стенки грамотрицательных бактерийУ грамотрицательных бактерий поверх клеточной мембраны тоже залегает слой пептидогликана, однако он значительно (почти в 40 раз[17]) тоньше, чем у грамположительных бактерий, и прикрыт сверху второй мембраной. Клеточная и наружная мембраны отличаются по химическому составу. Пространство между клеточной и наружной мембранами называется периплазматическим пространством (периплазмой)[27].
В периплазматическом пространстве находится много разнообразных белков: разрушающие биологические молекулы ферменты, транспортные белки, белки, участвующие в метаболизме, а также шапероны, которые регулируют пространственную структуру других белков, защищают их от протеолиза и нежелательных взаимодействий с другими белками. Например, в периплазме происходит образование дисульфидных мостиков и цис—транс-изомеризация пролина, которая является частью процесса созревания белка. Некоторые шапероны периплазмы участвуют в сборке ворсинок. Если под действием стрессовых условий в периплазме происходит агрегация[en] неуложенных белковых молекул, то активируется система Cpx. Она состоит из белка CpxA, заякоренного в клеточной мембране, и связанного с ним белка CpxP, который обращён в периплазму. CpxP взаимодействует с неуложенными белками и покидает CpxA, который при этом подвергается аутофосфорилированию[en] и далее переносит фосфатную группу на цитоплазматический белок CpxR. Фосфорилированный CpxR запускает экспрессию генов стрессового ответа[28]. По мере роста клетки в периплазматическом пространстве накапливаются продукты метаболизма пептидогликана, которые клетка использует повторно[29].
 Строение липополисахарида
Строение липополисахаридаНаружная мембрана состоит из двух асимметричных слоёв: внутренний слой, обращённый к клетке, состоит из фосфолипидов, а внешний — из липополисахаридов. Внутренний слой почти на 90 % состоит из фосфатидилэтаноламина[13]. Липополисахариды содержат О-полисахарид, коровый полисахарид и остаток липида А[en]. О-полисахарид, как правило, состоит из повторяющихся остатков галактозы, глюкозы, рамнозы и маннозы. Центральный (коровый) полисахарид состоит из N-ацетилглюкозамина, глюкозамина, фосфата, гептозы и кетодезоксиоктоната. Липополисахариды токсичны для животных и являются важнейшими антигенами, активирующими иммунную систему в ответ на бактериальное заражение[30]. Наружная мембрана связана со слоем пептидогликана при помощи липопротеинов, N-концы которых связаны с жирными кислотами и погружены во внешнюю мембрану, а C-концы связаны с пептидогликаном. Во внешней мембране имеются белки-порины[en], а также белки, связанные со сборкой поверхностных структур, конъюгацией и секрецией белковых молекул[31].
От наружной мембраны могут образовываться так называемые везикулы наружной мембраны, имеющие диаметр от 20 до 500 нм. В отпочковывании везикул принимает участие цитоскелет. Образование везикул может быть вызвано тем, что при росте клетки наружная мембрана увеличивается быстрее пептидогликанового слоя, а может вызываться особыми внешними условиями, например, у Porphyromonas gingivalis[en] образование везикул запускается нехваткой гемина[en]. Стенка везикул состоит из наружной мембраны, и при отпочковывании она может захватывать содержимое периплазмы. Так, у Pseudomonas aeruginosa везикулы наружной мембраны содержат периплазматические ферменты, в числе которых гемолизин, пептидогликан-гидролазы, протеазы, проэластазы, щелочная фосфатаза и фосфолипаза C, а также β-лактамаза, которая позволяет бактериям гидролизовать β-лактамные антибиотики и в периплазме, и во внешней среде. Везикулы наружной мембраны могут также служить для доставки ферментов и фрагмента наружной мембраны к клеткам-мишеням или в необходимый участок внешней среды[32].
В клетке грамотрицательной бактерии имеется от 200 до 400 зон слипания между наружной и клеточной мембранами, которые называют контактами Байера. В области контактов Байера в пептидогликановом слое имеется крупное отверстие, благодаря чему наружная и клеточная мембраны могут сблизиться вплотную. Адгезию мембран могут обеспечивать компоненты некоторых экспортных комплексов. Контакты Байера могут служить для выделения наружу различных молекул, например, субъединиц пилей, кроме того, к ним прикрепляются некоторые бактериофаги[33].
Из-за наличия дополнительного барьера проницаемости (наружной мембраны) для достижения необходимого эффекта грамотрицательные бактерии требуют больших концентраций антибиотиков, чем грамположительные бактерии. Наружная мембрана обеспечивает взаимодействия клеток друг с другом, с клетками организма-хозяина (при патогенезе) и с поверхностью субстрата. Она удерживает такие внешние структурные образования, как пили[29].
У цианобактерий поверх слоя пептидогликана располагается внешняя мембрана, однако с пептидогликаном ковалентно связаны полисахариды, из-за которых, судя по всему, цианобактерии окрашиваются по Граму положительно. Кроме того, наружная мембрана цианобактерий содержит каротиноиды[21].
Формирование клеточной стенки[править | править код]
Синтез пептидогликана протекает в несколько этапов. Дисахариды его гликановых цепей синтезируются в цитозоле, начиная с уридин-5-дифосфата[en]-NAM (УДФ-NAM). Синтез пептида начинается на NAM. Он формируется последовательно, присоединение каждой аминокислоты катализируется определённым ферментом. Наконец, пентапептид, связанный с УДФ-NAM, присоединяется к особому липиду клеточной мембраны — бактопренолу[en]. Далее к УДФ-NAM присоединяется NAG, УДФ высвобождается, и всё звено, включающее NAM, NAG и пептид, переворачивается и становится обращённым во внешнюю среду, а не в цитоплазму. Поперечные сшивки образуются с участием ферментов DD-транспептидаз[en] (которые ингибируются пенициллином), которые катализируют реакцию транспептидации, сопровождающуюся высвобождением остатка D-аланина. Образование гликановых цепей катализируют трансгликозилазы, кроме того, имеются бифункциональные ферменты, обладающие и трансгликозилазной, и транспептидазной активностями[19].
Согласно одной из предложенных моделей, при росте клетки в клеточную стенку сначала добавляются новые нити, и лишь потом происходит разрыв старых связей. У большинства бактерий в клеточной стенке находится множество ферментов автолиза, которые разрушают различные химические связи в пептидогликане[34].
Под системами секреции у бактерий понимают белковые комплексы, расположенные в клеточной мембране бактерий и служащие для выведения во внешнюю среду различных веществ. В частности, их используют патогенные бактерии для выделения факторов вирулентности[en] (преимущественно белковой природы). На основании состава, структура и действия системы секреции делят на несколько типов. Существует по меньшей мере шесть типов систем секреции, специфичных для грамотрицательных бактерий, четыре типа систем секреции уникальны для грамположительных бактерий, а два типа систем секреции имеются у обеих групп бактерий. Типы бактериальных систем секреции и их основные свойства приведены в таблице ниже[25].
| Система секреции | Сигнал секреции | Количество этапов секреции | Уложен ли субстрат | Количество мембран | Грам(+) или грам(-) |
|---|---|---|---|---|---|
| Sec | N-концевой | 1 | Нет | 1 | Обе группы |
| Tat | N-концевой | 1 | Да | 1 | Обе группы |
| I тип | C-концевой | 1 | Нет | 2 | Грам(-) |
| II тип | N-концевой | 2 | Да | 1 | Грам(-) |
| III тип | N-концевой | 1—2 | Нет | 2—3 | Грам(-) |
| IV тип | C-концевой | 1 | Нет | 2—3 | Грам(-) |
| V тип | N-концевой | 2 | Нет | 1 | Грам(-) |
| VI тип | Неизвестен | 1 | Неизвестно | 2—3 | Грам(-) |
| SecA2 | N-концевой | 1 | Нет | 1 | Грам(+) |
| Сортазы | N-концевой (Sec) C-концевой (cws) | 2 | Да | 1 | Грам(+) |
| Инжектосомы | N-концевой | 2 | Да | 1 | Грам(+) |
| VII тип | C-концевой | 1 | Да | 1—3 | Грам(+) |
Капсула[править | править код]
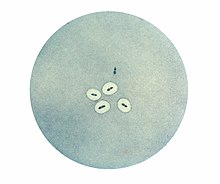 Клетки Streptococcus pneumoniae с капсулами, визуализированными при помощи Quellung-реакции. Обратите внимание, что две бактерии в верхней части фотографии не имеют капсулы
Клетки Streptococcus pneumoniae с капсулами, визуализированными при помощи Quellung-реакции. Обратите внимание, что две бактерии в верхней части фотографии не имеют капсулыУ многих бактерий поверх клеточной стенки или внешней мембраны залегает капсула, состоящая из экзополисахаридов[en]. Структурной основой капсулы служат линейные или разветвлённые полигликаны и олипептиды, состоящие из одинаковых или разных мономеров. У непатогенных бактерий капсулы служат для защиты высыхания, например, именно благодаря капсуле цианобактерии рода Nostoc могут расти в пустыне. У патогенных бактерий капсула резко увеличивает вирулентность, так как иммунная система плохо справляется с бактериями, покрытыми капсулой: они плохо связываются с антителами и не поддаются фагоцитированию[35].
S-слой[править | править код]
Поверхность некоторых бактерий (как грамположительных, так и грамотрицательных) покрыта S-слоем, состоящим из упорядоченно уложенных белковых субъединиц. У бактерий очень редки случаи, когда S-слой является единственной плотной оболочкой, обычно он сосуществует вместе с пептидогликановой клеточной стенкой. S-слой не играет формообразующей роли и часто утрачивается бактериями, растущими в лабораторных условиях. Сборка S-слоя начинается с того, что его белковые субъединицы секретируются в экзоплазматический компартмент, где они спонтанно агрегируют, связываясь друг с другом гидрофобными, водородными и электростатическими связями. S-слой обеспечивает механическую защиту бактериальной клетки, препятствует попаданию в клетку экзогенных молекул, взаимодействует с бактериофагами. У патогенных бактерий S-слой является важным фактором вирулентности[36].
Жгутик[править | править код]
 Схема строения бактериального жгутика
Схема строения бактериального жгутикаБольшинство бактерий подвижны, и их подвижность обеспечивается одним или нескольким жгутиками, которые представляют собой поверхностные белковые структуры. Расположение жгутиков на клетке может быть различным. У монотрихов имеется только один жгутик, у лофотрихов на одном из полюсов клетки находится пучок жгутиков, у амфитрихов на противоположных полюсах клетки находится по одному жгутику, а у перитрихов многочисленные жгутики разбросаны по всей поверхности клетки. Длина жгутика варьирует, но диаметр обычно составляет 20 нм[37].
Основание бактериального жгутика представлено базальным телом, состоящим из двух (у грамположительных) или четырёх (у грамотрицательных бактерий) белковых колец, стержня и моторных белков. От базального тела отходит крючок, переходящий в , который завершается «шапочкой». Филамент представляет собой жёсткий цилиндр, образованный белком флагеллином. В клеточной мембране находятся кольца M и S, которые часто рассматривают как единое целое. MS-кольцо окружено несколькими моторными белками, которые передают вращающий момент на филамент. У грамотрицательных бактерий, помимо колец M и S, есть ещё два кольца: P, залегающее в пептидогликановом слое, и L, находящееся во внешней мембране. Через все кольца проходит жёсткий стержень, передающий вращающий момент на филамент[38].
При сборке жгутика сначала в мембране клетки появляется MS-кольцо, к которому прикрепляются моторные белки, далее формируются P- и L-кольца (у грамотрицательных бактерий), крючок и филамент. В такой же последовательности запускается экспрессия генов, кодирующих белки соответствующей части жгутика[39]. Через полое внутреннее пространство базального тела новые флагеллиновые субъединицы поступают к вершине растущего жгутика, где самоорганизуются по спирали. Чтобы субъединицы флагеллина не уходили во внешнюю среду, конец растущего филамента прикрыт «шапочкой», которая не даёт им покинуть жгутик. В среднем зрелый филамент состоит из около 20 тысяч молекул флагеллина, а белки жгутика кодируются более чем 30 генами[40].
Вероятно, движущей силой вращения жгутика у бактерий является протонный градиент. Поток протонов, проходящий через кольца M и S или между базальным телом и клеточной мембраной, запускает вращение жгутика[41].
Движение клетки происходит за счёт вращения жгутика по часовой стрелке или против неё. У монотрихов клетка медленно вращается в направлении, противоположном вращению жгутика. Если жгутик вращается по часовой стрелке, то клетка движется жгутиком вперёд, а если против, то клетка выталкивается жгутиком вперёд (то есть движется жгутиком назад). Некоторые бактерии, имеющие единственный жгутик, вращают его только по часовой стрелке, и, чтобы сменить направление движения, им нужно остановиться и переориентироваться. У перитрихов жгутики вращаются против часовой стрелки, и, если нужно сменить направление движения, клетка останавливается и совершает кувырок[41].
У некоторых бактерий рода Vibrio (в частности, Vibrio parahaemolyticus[en][42]) и некоторых протеобактерий, таких как Aeromonas, имеются две различные жгутиковые системы, белковые компоненты которых кодируются различными наборами генов, а для вращения используются разные ионные градиенты. Полярные жгутики, относящиеся к первой жгутиковой системе, присутствуют постоянно и обеспечивают подвижность в потоке жидкости, а боковые жгутики, относящиеся ко второй жгутиковой системе, экспрессируются только тогда, когда сопротивление окружающей жидкости так велико, что полярные жгутики не могут вращаться. Благодаря этому бактерии могут скользить по различным поверхностям и в вязкой жидкости[43][44][45][46][47][48].
Пили[править | править код]
Клетки E. coli с многочисленными пилямиПили (также известны как фимбрии или ворсинки) — нитевидные белковые структуры, расположенные на поверхности клеток многих бактерий. Размер пилей варьирует от долей мкм до более чем 20 мкм в длину и 2—11 нм в диаметре. Пили участвуют в передаче генетического материала между бактериальными клетками (конъюгация), прикреплении бактерий к субстрату и другим клеткам, отвечают за адаптацию организмов, служат местами прикрепления многих бактериофагов. Структурно пили могут быть от тонких нитевидных образований до толстых палочкообразных структур с осевыми отверстиями. Пили состоят из одного или нескольких типов спирально уложенных белковых молекул, которые называют пилинами[en] или фимбринами[49]. В образовании пилей, помимо самих белков-пилинов, участвуют дополнительные белки, способствующие правильной сборке. У грамотрицательных бактерий они должны пройти через клеточную мембрану, периплазматическое пространство и наружную мембрану[50].
Прочие внеклеточные структуры[править | править код]
Иногда слизистая структура окружает не отдельную клетку, как в случае капсулы, а скопление клеток, и тогда покровную слизистую структуру называют чехлом. Чехлы могут покрывать не только вегетативные клетки, но и другие варианты дифференцированных клеток, например, покоящиеся структуры (цисты, эндоспоры, гетероцисты). Чехлы имеются, например, у внутриклеточны